Agriculture
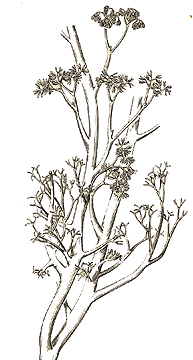 As first proposed by Harlan Banks in 1968, the trimerophytes evolved from the rhyniophytes and then gave rise, either directly or indirectly, to all other groups of vascular land plants except the zosterophyllophytes and lycopods.
As first proposed by Harlan Banks in 1968, the trimerophytes evolved from the rhyniophytes and then gave rise, either directly or indirectly, to all other groups of vascular land plants except the zosterophyllophytes and lycopods.
The trimerophytes appeared and diversified between 406 million and 401 million years ago, during the Devonian period. They evolved from the rhyniophytes (Rhyniophyta), and they share a number of characteristics with that group.
Both groups branched by having an axis fork into two branches of equal size. Viewed from the side, the point of branching would appear like a capital Y. Both groups also bore elongate sporangia at the ends of some of these branches.
The chief feature that distinguished the two groups was size. The rhyniophytes were small plants, approximately 25 centimeters (10 inches) or less in height.
The trimerophytes could reach heights in excess of 1 meter (1 yard). Because the trimerophytes were larger than the rhyniophytes, some species had a singlemain axis from which the lateral branches arose.
Establishing a taxonomic group just because some of its members were bigger than other contemporary plants is unusual. Some researchers believe that the group is too broadly defined to be taxonomically useful.
The two best-known genera, Psilophyton and Pertica, are more similar to early ferns and gymnosperms than they are to each other, supporting the belief than the trimerophytes are not a valid taxonomic group.
The smaller, herbaceous trimerophytes, such as Psilophyton, branched by forking into two branches of equal or unequal size and bore sporangia at the ends of some of these branches. Other trimerophytes, such as Pertica, were robust plants that probably reached the size of small shrubs.
They had a large, central axis that gave rise to smaller lateral branches. In Pertica quadrifaria and P. dalhousii, the lateral branches forked synchronously from four to six times before ending in either sterile tips or elongate sporangia. The sporangia opened (dehisced) along one side by means of a longitudinal slit.
Where known, the vascular tissue of the trimerophytes consisted entirely of primary phloem and xylem and seemed insufficient to support a plant more than a meter tall. The plants grew in dense clonal stands (a growth pattern called turfing), where the aerial axes could provide mutual support for each other.
At any site, about half of the trimerophytes present were fertile. The high proportion of fertile axes suggests plants that grew rapidly and reached reproductive maturity quickly. Since the sporangia all appear to be at the same stage of development, the aerial stems of the trimerophytes probably terminated their lives with a burst of reproduction.
New growth would then arise from the perennial rhizomes or root systems when favorable conditions returned. The trimerophytes preferred to live near freshwater in habitats thatwere susceptible to flooding. At this time in earth history, size was more important for spore dispersal than for light interception to power photosynthesis.
Trimerophyton robustius
The trimerophytes are named for Trimerophyton robustius, a Canadian plant originally described as Psilophyton robustius by J.W. Dawson. The genus is based on a single specimen about 12 centimeters (5 inches) long. Trimerophyton?s central axis gave rise to spirally arranged lateral branches. Initially, the lateral branches were believed to fork twice to produce a total of nine axes.
The first forking produced three new axes. The second forking of each newaxis produced three more axes for a total of nine. When viewed from the side, the point of branching appeared similar to a tripod. Each of these nine branches forked into two branches from two to three more times.
The ends of the branches were terminated by elongate sporangia that were all at the same stage of development. Reinterpretation of Dawson?s specimen indicates that the first divisions in the lateral branches resulted in two rather than three newaxes.
If this interpretation is correct, Trimerophyton no longer possesses the diagnostic trait that names and identifies the trimerophytes. The stemof Trimerophytonwas naked (lacked leaves or enations). Enations resemble leaves, but they lack vascular tissue and, therefore, have no veins. Both roots andrhizomes are unknown for Trimerophyton.
Psilophyton
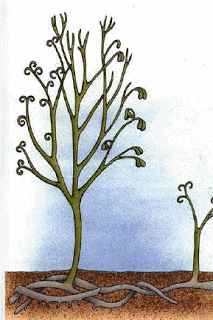 At least nine species of Psilophyton are known. They range in size from small plants (P. dapsile) that lacked a central main axis to larger plants that had prominent central axes. The stems could be naked (P. dapsile, P. dawsonii, and P. forbesii) or variously cloaked with spiny or peglike enations (P. crenulatum, P. princeps, and P. charientos).
At least nine species of Psilophyton are known. They range in size from small plants (P. dapsile) that lacked a central main axis to larger plants that had prominent central axes. The stems could be naked (P. dapsile, P. dawsonii, and P. forbesii) or variously cloaked with spiny or peglike enations (P. crenulatum, P. princeps, and P. charientos).
The short species and the lateral branch systems of the larger species branched by forking to produce two new axes. The sterile branches end in blunt tips and the fertile ones in paired sporangia. The sporangia occur in dense clusters of thirty-two or more.
Psilophyton princeps was the first valid species described. Dawson named his Canadian fossils in 1859 but did not publish a reconstruction of the plant until 1870. As reconstructed by Dawson, the naked aerial stems arose from a rhizome (a stem that runs horizontally along the ground) and branched by forking.
Sporangia were borne at the ends of the branches and hung down toward the ground (that is, the sporangia were pendant) Dawson also figured a spiny axis that he named P. princeps var. ornatum. This variety was subsequently found to have lateral sporangia and was redescribed as Sawdonia ornata. Sawdonia is classi fied as a zosterophyllophyte.
None of the parts (aerial stem, sporangia, or rhizome) of Dawson?s original Psilophyton princeps were found attached to each other. They were ultimately found to represent parts of three distinct plants.
Psilophyton princeps itself was covered with short, peglike enations and bore clusters of terminal sporangia at the ends of some of its lateral branches. The naked axis bearing the pendant sporangia was named Dawsonites arcuatus. Dawsonites has remained in the Trimerophytophyta for convenience because scientists are not sure of its exact affinities.
The rhizome was renamed Taeniocrada dubia and may belong in the Rhyniophyta (possibly in the genus Stockmansella). Taeniocrada is simply a long, naked axis that lacks any unique structural features.
Pertica
 Plants of this genus (P. quadrifolia, P. dalhousii, and P. varia) are the largest trimerophytes, reaching heights of ameter ormore. The stemappears naked but is actually covered with small (0.4-millimeter) bumps call papillae.
Plants of this genus (P. quadrifolia, P. dalhousii, and P. varia) are the largest trimerophytes, reaching heights of ameter ormore. The stemappears naked but is actually covered with small (0.4-millimeter) bumps call papillae.
The strongly developed main axis gave rise to short lateral branches in groups of four. The lateral branches could fork into two or three new axes of equal size.
These axes could be either sterile or fertile. The fertile branches were mixed in with the sterile branches. The fertile branches end in dense, spherical clusters of round to elongate sporangia, which opened by a slit down the side.
If Dawson?s specimen of Trimerophyton did produce some lateral branches by forking into three axes, the specimen may actually represent a short segment of P. varia that branched in this fashion.
Progeny
The greatmajority of both fossil and living plants can trace their lineages back to the trimerophytes, specifically to the Psilophyton-Pertica complex. Derived groups include the ferns, horsetails, gymnosperms, and angiosperms. The trimerophytes are very similar to the early ferns and gymnosperms, most notably to the extinct progymnosperm Tetraxylopteris.
The resemblance is so great that some researchers feel that the Trimerophytophyta is not a validgroup. The trimerophytes are very different in appearance from the ferns, gymnosperms, and angiosperms that are alive today.
The groups that evolved from the trimerophytes had far greater impact on the global environment than their predecessors, the rhyniophytes, zosterophyllophytes, and trimerophytes, did together. These latter groups had a very narrow habitat range, had shallow rooting systems or rhizomes, and were not seed producers.
Their successors had well-developed root systems, as seen in the increased thickness and horizontal zonation of fossil soils (paleosols). The development of seeds allowed the gymnosperms and angiosperms to escape from a dependence on moist, lowland habitats to ensure reproductive success and to colonize drier, upland habitats.
The development of seeds allowed the spread of forests from 377 million to 362 million years ago. The spread of forests was followed by a worldwide increase in the deposition of black shale and the formation of coal.
The organic material represented by these deposits reflected a significant loss of carbon dioxide from the atmosphere. The decrease in atmospheric carbon dioxide brought on a period of continental glaciation and caused a mass extinction of tropical marine invertebrates due to decreasedwater temperature.
The tropical sea?s surface temperate cooled from 40 degrees Celsius (104 degrees Fahrenheit) about 345 million years ago to between 24 and 26 degrees Celsius (77 degrees Fahrenheit) about 280 million years ago.
- Gymnosperms
GymnospermsPine trees are a familiar example of gymnosperms, a series of evolutionary lines of woody vascular seed plants that produce seeds not encased in an ovary. Two kinds of higher plants?the gymnosperms and angiosperms?have developed to become the...
- Lycophytes
LycophytesThe lycophytes, which compose the phylum Lycophyta, are one of four phyla of seedless plants having vascular, or conducting, tissue. The living lycophytes are all small and herbaceous, whereas the extinct lycophytes included large trees, which...
- Psilotophytes
PsilotophytesPsilotophyte is the common name for members of the phylum Psilotophyta (from the Greek word psilos, meaning ?bare?). Molecular evidence points to the likelihood of psilotophytes as being highly reduced (and therefore derived) ferns. If psilotophytes...
- Rhyniophyta
RhyniophytaWhen first proposed by Harlan Banks in 1968, the Rhyniophyta were the first and oldest vascular land plants. The trimerophytes subsequently evolved from them. In 1908, Octave Lignier developed a model of what the sporophyte of the earliest...
- Seedless Vascular Plants
Seedless Vascular Plants Seedless vascular plants possess vascular tissues (xylem and phloem) for transport of materials through the body but do not produce seeds bearing dormant embryos as part of the reproductive process. They are among the oldest of...
Agriculture
Trimerophytophyta
The trimerophytes appeared and diversified between 406 million and 401 million years ago, during the Devonian period. They evolved from the rhyniophytes (Rhyniophyta), and they share a number of characteristics with that group.
Both groups branched by having an axis fork into two branches of equal size. Viewed from the side, the point of branching would appear like a capital Y. Both groups also bore elongate sporangia at the ends of some of these branches.
The chief feature that distinguished the two groups was size. The rhyniophytes were small plants, approximately 25 centimeters (10 inches) or less in height.
The trimerophytes could reach heights in excess of 1 meter (1 yard). Because the trimerophytes were larger than the rhyniophytes, some species had a singlemain axis from which the lateral branches arose.
Establishing a taxonomic group just because some of its members were bigger than other contemporary plants is unusual. Some researchers believe that the group is too broadly defined to be taxonomically useful.
The two best-known genera, Psilophyton and Pertica, are more similar to early ferns and gymnosperms than they are to each other, supporting the belief than the trimerophytes are not a valid taxonomic group.
The smaller, herbaceous trimerophytes, such as Psilophyton, branched by forking into two branches of equal or unequal size and bore sporangia at the ends of some of these branches. Other trimerophytes, such as Pertica, were robust plants that probably reached the size of small shrubs.
They had a large, central axis that gave rise to smaller lateral branches. In Pertica quadrifaria and P. dalhousii, the lateral branches forked synchronously from four to six times before ending in either sterile tips or elongate sporangia. The sporangia opened (dehisced) along one side by means of a longitudinal slit.
Where known, the vascular tissue of the trimerophytes consisted entirely of primary phloem and xylem and seemed insufficient to support a plant more than a meter tall. The plants grew in dense clonal stands (a growth pattern called turfing), where the aerial axes could provide mutual support for each other.
At any site, about half of the trimerophytes present were fertile. The high proportion of fertile axes suggests plants that grew rapidly and reached reproductive maturity quickly. Since the sporangia all appear to be at the same stage of development, the aerial stems of the trimerophytes probably terminated their lives with a burst of reproduction.
New growth would then arise from the perennial rhizomes or root systems when favorable conditions returned. The trimerophytes preferred to live near freshwater in habitats thatwere susceptible to flooding. At this time in earth history, size was more important for spore dispersal than for light interception to power photosynthesis.
Trimerophyton robustius
The trimerophytes are named for Trimerophyton robustius, a Canadian plant originally described as Psilophyton robustius by J.W. Dawson. The genus is based on a single specimen about 12 centimeters (5 inches) long. Trimerophyton?s central axis gave rise to spirally arranged lateral branches. Initially, the lateral branches were believed to fork twice to produce a total of nine axes.
The first forking produced three new axes. The second forking of each newaxis produced three more axes for a total of nine. When viewed from the side, the point of branching appeared similar to a tripod. Each of these nine branches forked into two branches from two to three more times.
The ends of the branches were terminated by elongate sporangia that were all at the same stage of development. Reinterpretation of Dawson?s specimen indicates that the first divisions in the lateral branches resulted in two rather than three newaxes.
If this interpretation is correct, Trimerophyton no longer possesses the diagnostic trait that names and identifies the trimerophytes. The stemof Trimerophytonwas naked (lacked leaves or enations). Enations resemble leaves, but they lack vascular tissue and, therefore, have no veins. Both roots andrhizomes are unknown for Trimerophyton.
Psilophyton
The short species and the lateral branch systems of the larger species branched by forking to produce two new axes. The sterile branches end in blunt tips and the fertile ones in paired sporangia. The sporangia occur in dense clusters of thirty-two or more.
Psilophyton princeps was the first valid species described. Dawson named his Canadian fossils in 1859 but did not publish a reconstruction of the plant until 1870. As reconstructed by Dawson, the naked aerial stems arose from a rhizome (a stem that runs horizontally along the ground) and branched by forking.
Sporangia were borne at the ends of the branches and hung down toward the ground (that is, the sporangia were pendant) Dawson also figured a spiny axis that he named P. princeps var. ornatum. This variety was subsequently found to have lateral sporangia and was redescribed as Sawdonia ornata. Sawdonia is classi fied as a zosterophyllophyte.
None of the parts (aerial stem, sporangia, or rhizome) of Dawson?s original Psilophyton princeps were found attached to each other. They were ultimately found to represent parts of three distinct plants.
Psilophyton princeps itself was covered with short, peglike enations and bore clusters of terminal sporangia at the ends of some of its lateral branches. The naked axis bearing the pendant sporangia was named Dawsonites arcuatus. Dawsonites has remained in the Trimerophytophyta for convenience because scientists are not sure of its exact affinities.
The rhizome was renamed Taeniocrada dubia and may belong in the Rhyniophyta (possibly in the genus Stockmansella). Taeniocrada is simply a long, naked axis that lacks any unique structural features.
Pertica
The strongly developed main axis gave rise to short lateral branches in groups of four. The lateral branches could fork into two or three new axes of equal size.
These axes could be either sterile or fertile. The fertile branches were mixed in with the sterile branches. The fertile branches end in dense, spherical clusters of round to elongate sporangia, which opened by a slit down the side.
If Dawson?s specimen of Trimerophyton did produce some lateral branches by forking into three axes, the specimen may actually represent a short segment of P. varia that branched in this fashion.
Progeny
The greatmajority of both fossil and living plants can trace their lineages back to the trimerophytes, specifically to the Psilophyton-Pertica complex. Derived groups include the ferns, horsetails, gymnosperms, and angiosperms. The trimerophytes are very similar to the early ferns and gymnosperms, most notably to the extinct progymnosperm Tetraxylopteris.
The resemblance is so great that some researchers feel that the Trimerophytophyta is not a validgroup. The trimerophytes are very different in appearance from the ferns, gymnosperms, and angiosperms that are alive today.
The groups that evolved from the trimerophytes had far greater impact on the global environment than their predecessors, the rhyniophytes, zosterophyllophytes, and trimerophytes, did together. These latter groups had a very narrow habitat range, had shallow rooting systems or rhizomes, and were not seed producers.
Their successors had well-developed root systems, as seen in the increased thickness and horizontal zonation of fossil soils (paleosols). The development of seeds allowed the gymnosperms and angiosperms to escape from a dependence on moist, lowland habitats to ensure reproductive success and to colonize drier, upland habitats.
The development of seeds allowed the spread of forests from 377 million to 362 million years ago. The spread of forests was followed by a worldwide increase in the deposition of black shale and the formation of coal.
The organic material represented by these deposits reflected a significant loss of carbon dioxide from the atmosphere. The decrease in atmospheric carbon dioxide brought on a period of continental glaciation and caused a mass extinction of tropical marine invertebrates due to decreasedwater temperature.
The tropical sea?s surface temperate cooled from 40 degrees Celsius (104 degrees Fahrenheit) about 345 million years ago to between 24 and 26 degrees Celsius (77 degrees Fahrenheit) about 280 million years ago.
- Gymnosperms
GymnospermsPine trees are a familiar example of gymnosperms, a series of evolutionary lines of woody vascular seed plants that produce seeds not encased in an ovary. Two kinds of higher plants?the gymnosperms and angiosperms?have developed to become the...
- Lycophytes
LycophytesThe lycophytes, which compose the phylum Lycophyta, are one of four phyla of seedless plants having vascular, or conducting, tissue. The living lycophytes are all small and herbaceous, whereas the extinct lycophytes included large trees, which...
- Psilotophytes
PsilotophytesPsilotophyte is the common name for members of the phylum Psilotophyta (from the Greek word psilos, meaning ?bare?). Molecular evidence points to the likelihood of psilotophytes as being highly reduced (and therefore derived) ferns. If psilotophytes...
- Rhyniophyta
RhyniophytaWhen first proposed by Harlan Banks in 1968, the Rhyniophyta were the first and oldest vascular land plants. The trimerophytes subsequently evolved from them. In 1908, Octave Lignier developed a model of what the sporophyte of the earliest...
- Seedless Vascular Plants
Seedless Vascular Plants Seedless vascular plants possess vascular tissues (xylem and phloem) for transport of materials through the body but do not produce seeds bearing dormant embryos as part of the reproductive process. They are among the oldest of...