Agriculture
Mendelian genetics is the classical mechanistic explanation of heredity in sexually reproducing organisms. It explains how genetic information is passed from one generation to another.
In 1866 the Augustinian monk Gregor Mendel (1822-1884) published a paper titled ?Versuch über Pflanzenhybriden? (Experiments in Plant Hybridisation, 1910), describing the heredity of mutant characteristics of garden peas.
Mendel founded the modern science of genetics with these experiments, because they led him to propose the existence of hereditary factors, now called genes, and rules describing their inheritance, now referred to as Mendel?s laws. The importance of Mendel?s work was not recognized until 1900, sixteen years after his death, when the movements of chromosomes during cell division were carefully studied.
Since then, Mendel?s laws have been shown to hold true throughout nature. The biochemical nature of genes has been discovered, the genetic code has been broken, and genetics has assumed a central role in modern biology, medicine, and agriculture.
Mendel?s Experiments
Mendel was not the only researcher interested in genetics, or inheritance, as it was called in his day. The prevailing theories of his time, though, differed considerably from his final conclusions. It was believed by most biologists that inheritance involved the blending of some sort of element from each parent. The result is offspring that are intermediate between the parents.
This process, as understood, was analogous to blending two colors of paint. Experiments by others made it difficult to challenge this theory, because they often studied numerous complex traits simultaneously, only analyzed offspring from a single generation, and worked with small numbers of organisms.
One of the reasons for Mendel?s success was that he simplified the problem of heredity by analyzing a few simple, easily distinguishable, hereditary differences among a species that was easy to breed. He also initially studied one trait at a time and followed the inheritance of each trait for several generations, using large enough numbers to solidify his conclusions.
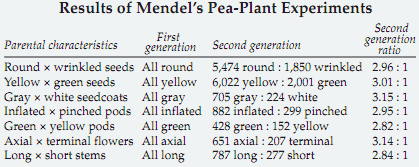
He began by selecting strains of garden peas that differed by a single trait from normal strains, such as wrinkled versus smooth, green peas versus yellow, and tall plants versus short. Once each strain bred true for a mutant variation, he crossed it with a different strain to see which trait was passed on to the offspring.
Mendel observed that all the hybrid offspring of each individual cross resembled one of the parent types and not the other, rather than a mixture of the two types or an intermediate form (the conventional ?mixed-paint? theory). Crosses between tall and short parent strains, for example, produced hybrids that were tall only. Mendel defined this phenomenon as dominance of one trait over the alternate trait, which he called the recessive trait.
Mendel then discovered that crossing two hybrids resulted in the reappearance of the recessive trait but only in one-fourth of the offspring. A cross between two tall hybrids, for example, produced about three-fourths tall plants and one fourth short plants. Mendel proposed that hereditary factors (now called genes) existed for each of the traits with which he was working.
He also proposed that hereditary factors exist in pairs, such that each individual inherits one from the pair carried by each parent. Mendel hypothesized that the pairs of factors would be separated, and one would be randomly included in each gamete (male or female germ cell?pollen or ovule, in the case of plants).
Testing the Theory
Mendel tested this theory with further crosses. (Because Mendel did not know of the existence of genes, he did not have a clearway to refer to the genetic factors responsible for his results. The word ?gene? is used in this discussion for convenience.)
He predicted, for example, that the original tall parent had two tall genes for height (symbolized as TT), and the original short parent had two short genes for height (tt). The hybrid would inherit one of each (Tt), but because the tall gene is dominant, the hybrid?s appearancewould be tall. Mendel predicted that crossing a Tt hybrid with one of the short (tt) plants should produce half tall (Tt)off-spring and half short ones (tt).
That is exactly what Mendel observed. He was also able to predict the outcomes of other crosses involving different traits. Mendel concluded that his theory worked: Paired hereditary factors must exist, and only one of the two, chosen at random, could be passed on to each offspring by each parent.
Mendel labeled this phenomenon segregation, meaning that the parent?s two hereditary factors are physically segregated into different cells during the production of gametes. This principle of segregation is now called Mendel?s first law of inheritance.
Mendel?s second law of inheritance describes the principle of independent assortment, which states that different hereditary factors segregate independently of one another. Mendel observed, for example, that if he crossed a tall and purple parent (TT-PP) with a short and white one (tt-pp), the hybrid offspring were tall and purple, as these genes are dominant.
Then, when he crossed the tall and purple hybrid (Tt-Pp) with another, identical hybrid, the progeny showed an ?assortment? of the two traits (tall and purple, tall and white, short and purple, short and white) in a 9:3:3:1 ratio, respectively.
This is the ratio expected if each trait?s genes segregate independently. Stated another way, whether a parent passes on a tall or short factor does not influence whether that parent also passes on the purple or white factor.
These two laws of heredity summarize Mendel?s discovery of discrete genetic factors and their patterns of inheritance: Mendel had proposed that discrete genetic factors exist, had explained how they are passed on, and had supported his theories with experimental evidence.
Mendel?s discovery, however, was virtually ignored. He died in 1884 without receiving recognition for his work. Mendel?s laws were independently rediscovered in the year 1900, and then their fundamental importance and general applicability were widely recognized.
Morgan?s Contributions
Microscopic bodies in the nuclei of cells, called chromosomes, had been discovered by the end of the nineteenth century, and in 1901 it was proposed that chromosomes are the physical structures that contain Mendel?s hereditary factors, or genes.
Chromosomes were a likely structure for the location of genes because chromosomes occur in pairs, duplicate when the cell divides, and segregate into sperm and egg cells such that only one of the two chromosomes in each pair is passed on to any single offspring by each parent. The chromosomal theory of heredity made it easier for biologists to think of genes as physical objects of analysis, and studies of Mendelian patterns of inheritance and their chromosomal basis progressed rapidly.
Ageneticist named Thomas Hunt Morgan at Columbia University made several key discoveries using fruit flies between 1910 and 1920. He and his colleagues discovered mutations in flies that showed different patterns of inheritance in males and females, which led to association of these genes with the sex-determining X and Y chromosomes.
Traits affected by genes on these chromosomes show a sex-linked pattern of inheritance in which recessive traits appear more often in males than in females. Human sex-linked traits, for example, include hemophilia, color-blindness, and baldness.
Fruit flies have three pairs of chromosomes besides the sex chromosomes, and Morgan?s laboratory team showed that traits could be grouped together in ?linkage groups? corresponding to their four pairs of chromosomes. They realized that Mendel?s second law describing the principle of independent assortment corresponded to the assortment of chromosomes being passed from parent to offspring.
Any genes on different chromosomes would be passed on independently, while genes linked together on the same chromosome would be passed on together as a unit. The discovery of linkage groups supported the idea that chromosomes were made up of collections of a large number of genes linked together.
Sturtevant?s Contributions
Morgan?s laboratory group, however, also observed occasional exceptions to this pattern of linkage,when offspring showed unexpected new combinations of linked genes that did not exist in either parent.
Alfred H. Sturtevant, a student in Morgan?s laboratory, proposed that the paired chromosomes carrying different forms of the same genes (one carrying recessive forms, for example, a-b-c, versus the other, carrying dominant forms A-B-C) could undergo a reciprocal exchange of part of the chromosome. One chromosome pair could exchange, for example, C for c, resulting in new a-b-C and A-B-c combinations of the linked genes.
Sturtevant also discovered that such recombination events occur with different frequencies between different genes. Only 1 percent of the A and B genes might be switched in each cross, for example, but 20 percent of the A and C genes might recombine in the same cross. Sturtevant proposed that the genes are linked together in a linear sequence and that the frequency of recombination between them is a function of the physical distance separating them on the chromosome.
Two genes that are far apart should recombine more frequently than two genes close together, since there would be a greater opportunity for the breakage and the exchange of different chromosomal material to occur between them.
Sturtevant proposed that differences in the frequency of recombination among linked genes on the same chromosome could be used to ?map? the genes in a linear sequence that would reveal their order and relative positions on the chromosome.
This principle turned out to be universal, and it allows genes to be mapped to specific locations on each chromosome in all organisms that can be systematically bred. Mendel?s genes had, by the 1920?s, been associated with chromosomes, and individual genes on each chromosome could be ordered and mapped using recombination analysis.
Mid-Twentieth Century Developments
The following two decades were marked by two important parallel developments in genetics. The first was a mathematical and experimental synthesis of Mendel?s genetic theory with Charles Darwin?s theory of natural selection.
It was shown that the genetic mechanism described by Mendel provided the hereditary mechanism required for Darwin?s theory of natural selection. The revision of Darwin?s work that resulted is often referred to as the neo-Darwinian synthesis.
The second development was progress in identifying the biochemical nature of genes, primarily by the extension of genetic analysis to bacteria and viruses. These studies led to the identification of deoxyribonucleic acid (DNA) as the hereditary molecule and to the identification of its biochemical structure by James Watson and Francis Crick in 1953.
Once the biochemical structure of genes was identified, an understanding of how DNA replicates and carries a genetic code that directs the synthesis of proteins followed rapidly.
One more revolutionary breakthrough that set the stage for the current era of genetics was the rapid development of recombinant DNA technology in the 1970?s and its refinement and broad application in the 1980?s. Recombinant DNA technology is a collection of methods that allows DNA sequences of one organism to be recombined with those of another. The application of these techniques is commonly referred to as ?genetic engineering.?
The fact that the chemical structure of DNA and the genetic code for protein synthesis are virtually the same for all organisms makes recombinant DNA a powerful technology. Recombinant DNA techniques, together with an understanding of the genetic code and the ability to identify and map specific genes, have opened up a new era of biological investigation and applications to medicine and agriculture.
- Chloroplast Dna
Chloroplast DNAPlants are unique among higher organisms in that they meet their energy needs through photosynthesis. The specific location for photosynthesis in plant cells is the chloroplast, which also contains a single, circular chromosome composed...
- Extranuclear Inheritance
Extranuclear Inheritance Extranuclear inheritance is a non-Mendelian form of heredity that involves genetic information located in cytoplasmic organelles, such as mitochondria and chloroplasts, rather than on the chromosomes found in the cell nucleus....
- Genetic Equilibrium: Linkage
Genetic Equilibrium: Linkage Genetic equilibriumis the tendency for genes located close together on the same chromosome to be inherited together. The farther apart the genes, the less likely it is that they will be passed along together. The genetic complement...
- Hybridization
HybridizationHybridization is the process of crossing two genetically different individuals to result in a third individual with a different, often preferred, set of traits. Plants of the same species cross easily and produce fertile progeny. Wide crosses...
- It Skips A Generation
Long before they understood why the strategy worked, farmers knew how to crossbreed plants to obtain more desirable traits. Even today, a farmer who knows nothing about genetics can tell you that when a blue type of corn crosses with a yellow one, the...
Agriculture
Genetics: Mendelian
") |
| Gregor Mendel (1822-1884) |
In 1866 the Augustinian monk Gregor Mendel (1822-1884) published a paper titled ?Versuch über Pflanzenhybriden? (Experiments in Plant Hybridisation, 1910), describing the heredity of mutant characteristics of garden peas.
Mendel founded the modern science of genetics with these experiments, because they led him to propose the existence of hereditary factors, now called genes, and rules describing their inheritance, now referred to as Mendel?s laws. The importance of Mendel?s work was not recognized until 1900, sixteen years after his death, when the movements of chromosomes during cell division were carefully studied.
Since then, Mendel?s laws have been shown to hold true throughout nature. The biochemical nature of genes has been discovered, the genetic code has been broken, and genetics has assumed a central role in modern biology, medicine, and agriculture.
Mendel?s Experiments
 |
| Mendel?s Experiments |
This process, as understood, was analogous to blending two colors of paint. Experiments by others made it difficult to challenge this theory, because they often studied numerous complex traits simultaneously, only analyzed offspring from a single generation, and worked with small numbers of organisms.
One of the reasons for Mendel?s success was that he simplified the problem of heredity by analyzing a few simple, easily distinguishable, hereditary differences among a species that was easy to breed. He also initially studied one trait at a time and followed the inheritance of each trait for several generations, using large enough numbers to solidify his conclusions.
He began by selecting strains of garden peas that differed by a single trait from normal strains, such as wrinkled versus smooth, green peas versus yellow, and tall plants versus short. Once each strain bred true for a mutant variation, he crossed it with a different strain to see which trait was passed on to the offspring.
Mendel observed that all the hybrid offspring of each individual cross resembled one of the parent types and not the other, rather than a mixture of the two types or an intermediate form (the conventional ?mixed-paint? theory). Crosses between tall and short parent strains, for example, produced hybrids that were tall only. Mendel defined this phenomenon as dominance of one trait over the alternate trait, which he called the recessive trait.
Mendel then discovered that crossing two hybrids resulted in the reappearance of the recessive trait but only in one-fourth of the offspring. A cross between two tall hybrids, for example, produced about three-fourths tall plants and one fourth short plants. Mendel proposed that hereditary factors (now called genes) existed for each of the traits with which he was working.
He also proposed that hereditary factors exist in pairs, such that each individual inherits one from the pair carried by each parent. Mendel hypothesized that the pairs of factors would be separated, and one would be randomly included in each gamete (male or female germ cell?pollen or ovule, in the case of plants).
Testing the Theory
Mendel tested this theory with further crosses. (Because Mendel did not know of the existence of genes, he did not have a clearway to refer to the genetic factors responsible for his results. The word ?gene? is used in this discussion for convenience.)
He predicted, for example, that the original tall parent had two tall genes for height (symbolized as TT), and the original short parent had two short genes for height (tt). The hybrid would inherit one of each (Tt), but because the tall gene is dominant, the hybrid?s appearancewould be tall. Mendel predicted that crossing a Tt hybrid with one of the short (tt) plants should produce half tall (Tt)off-spring and half short ones (tt).
That is exactly what Mendel observed. He was also able to predict the outcomes of other crosses involving different traits. Mendel concluded that his theory worked: Paired hereditary factors must exist, and only one of the two, chosen at random, could be passed on to each offspring by each parent.
Mendel labeled this phenomenon segregation, meaning that the parent?s two hereditary factors are physically segregated into different cells during the production of gametes. This principle of segregation is now called Mendel?s first law of inheritance.
 |
 |


Mendel?s second law of inheritance describes the principle of independent assortment, which states that different hereditary factors segregate independently of one another. Mendel observed, for example, that if he crossed a tall and purple parent (TT-PP) with a short and white one (tt-pp), the hybrid offspring were tall and purple, as these genes are dominant.
Then, when he crossed the tall and purple hybrid (Tt-Pp) with another, identical hybrid, the progeny showed an ?assortment? of the two traits (tall and purple, tall and white, short and purple, short and white) in a 9:3:3:1 ratio, respectively.
This is the ratio expected if each trait?s genes segregate independently. Stated another way, whether a parent passes on a tall or short factor does not influence whether that parent also passes on the purple or white factor.
These two laws of heredity summarize Mendel?s discovery of discrete genetic factors and their patterns of inheritance: Mendel had proposed that discrete genetic factors exist, had explained how they are passed on, and had supported his theories with experimental evidence.
Mendel?s discovery, however, was virtually ignored. He died in 1884 without receiving recognition for his work. Mendel?s laws were independently rediscovered in the year 1900, and then their fundamental importance and general applicability were widely recognized.
Morgan?s Contributions
 |
| green peas |
Chromosomes were a likely structure for the location of genes because chromosomes occur in pairs, duplicate when the cell divides, and segregate into sperm and egg cells such that only one of the two chromosomes in each pair is passed on to any single offspring by each parent. The chromosomal theory of heredity made it easier for biologists to think of genes as physical objects of analysis, and studies of Mendelian patterns of inheritance and their chromosomal basis progressed rapidly.
Ageneticist named Thomas Hunt Morgan at Columbia University made several key discoveries using fruit flies between 1910 and 1920. He and his colleagues discovered mutations in flies that showed different patterns of inheritance in males and females, which led to association of these genes with the sex-determining X and Y chromosomes.
Traits affected by genes on these chromosomes show a sex-linked pattern of inheritance in which recessive traits appear more often in males than in females. Human sex-linked traits, for example, include hemophilia, color-blindness, and baldness.
Fruit flies have three pairs of chromosomes besides the sex chromosomes, and Morgan?s laboratory team showed that traits could be grouped together in ?linkage groups? corresponding to their four pairs of chromosomes. They realized that Mendel?s second law describing the principle of independent assortment corresponded to the assortment of chromosomes being passed from parent to offspring.
Any genes on different chromosomes would be passed on independently, while genes linked together on the same chromosome would be passed on together as a unit. The discovery of linkage groups supported the idea that chromosomes were made up of collections of a large number of genes linked together.
Sturtevant?s Contributions
Morgan?s laboratory group, however, also observed occasional exceptions to this pattern of linkage,when offspring showed unexpected new combinations of linked genes that did not exist in either parent.
Alfred H. Sturtevant, a student in Morgan?s laboratory, proposed that the paired chromosomes carrying different forms of the same genes (one carrying recessive forms, for example, a-b-c, versus the other, carrying dominant forms A-B-C) could undergo a reciprocal exchange of part of the chromosome. One chromosome pair could exchange, for example, C for c, resulting in new a-b-C and A-B-c combinations of the linked genes.
Sturtevant also discovered that such recombination events occur with different frequencies between different genes. Only 1 percent of the A and B genes might be switched in each cross, for example, but 20 percent of the A and C genes might recombine in the same cross. Sturtevant proposed that the genes are linked together in a linear sequence and that the frequency of recombination between them is a function of the physical distance separating them on the chromosome.
Two genes that are far apart should recombine more frequently than two genes close together, since there would be a greater opportunity for the breakage and the exchange of different chromosomal material to occur between them.
Sturtevant proposed that differences in the frequency of recombination among linked genes on the same chromosome could be used to ?map? the genes in a linear sequence that would reveal their order and relative positions on the chromosome.
This principle turned out to be universal, and it allows genes to be mapped to specific locations on each chromosome in all organisms that can be systematically bred. Mendel?s genes had, by the 1920?s, been associated with chromosomes, and individual genes on each chromosome could be ordered and mapped using recombination analysis.
Mid-Twentieth Century Developments
 |
| How to grow juicy tasty tomatoes |
It was shown that the genetic mechanism described by Mendel provided the hereditary mechanism required for Darwin?s theory of natural selection. The revision of Darwin?s work that resulted is often referred to as the neo-Darwinian synthesis.
The second development was progress in identifying the biochemical nature of genes, primarily by the extension of genetic analysis to bacteria and viruses. These studies led to the identification of deoxyribonucleic acid (DNA) as the hereditary molecule and to the identification of its biochemical structure by James Watson and Francis Crick in 1953.
Once the biochemical structure of genes was identified, an understanding of how DNA replicates and carries a genetic code that directs the synthesis of proteins followed rapidly.
One more revolutionary breakthrough that set the stage for the current era of genetics was the rapid development of recombinant DNA technology in the 1970?s and its refinement and broad application in the 1980?s. Recombinant DNA technology is a collection of methods that allows DNA sequences of one organism to be recombined with those of another. The application of these techniques is commonly referred to as ?genetic engineering.?
The fact that the chemical structure of DNA and the genetic code for protein synthesis are virtually the same for all organisms makes recombinant DNA a powerful technology. Recombinant DNA techniques, together with an understanding of the genetic code and the ability to identify and map specific genes, have opened up a new era of biological investigation and applications to medicine and agriculture.
- Chloroplast Dna
Chloroplast DNAPlants are unique among higher organisms in that they meet their energy needs through photosynthesis. The specific location for photosynthesis in plant cells is the chloroplast, which also contains a single, circular chromosome composed...
- Extranuclear Inheritance
Extranuclear Inheritance Extranuclear inheritance is a non-Mendelian form of heredity that involves genetic information located in cytoplasmic organelles, such as mitochondria and chloroplasts, rather than on the chromosomes found in the cell nucleus....
- Genetic Equilibrium: Linkage
Genetic Equilibrium: Linkage Genetic equilibriumis the tendency for genes located close together on the same chromosome to be inherited together. The farther apart the genes, the less likely it is that they will be passed along together. The genetic complement...
- Hybridization
HybridizationHybridization is the process of crossing two genetically different individuals to result in a third individual with a different, often preferred, set of traits. Plants of the same species cross easily and produce fertile progeny. Wide crosses...
- It Skips A Generation
Long before they understood why the strategy worked, farmers knew how to crossbreed plants to obtain more desirable traits. Even today, a farmer who knows nothing about genetics can tell you that when a blue type of corn crosses with a yellow one, the...