Agriculture
Alternative forms of photosynthesis are used by specific types of plants, called C4 and CAM plants, to alleviate problems of photorespiration and excess water loss.
Photosynthesis is the physiological process whereby plants use the sun?s radiant energy to produce organic molecules. The backbone of all such organic compounds is a skeleton composed of carbon atoms. Plants use carbon dioxide from the atmosphere as their carbon source.
The overwhelming majority of plants use a single chemical reaction to attach carbon dioxide from the atmosphere onto an organic compound, a process referred to as carbon fixation. This process takes place inside specialized structures within the cells of green plants known as chloroplasts.
The enzyme that catalyzes this fixation is ribulose bisphosphate carboxylase (Rubisco), and the first stable organic product is a three-carbon molecule. This three-carbon compound is involved in the biochemical pathway known as the Calvin cycle. Plants using carbon fixation are referred to as C3 plants because the first product made with carbon dioxide is a three-carbon molecule.
C4 Photosynthesis
For many years scientists thought that the only way photosynthesis occurred was through C3 photosynthesis. In the early 1960?s, however, researchers studying the sugarcane plant discovered a biochemical pathway that involved incorporation of carbon dioxide into organic products at two different stages.
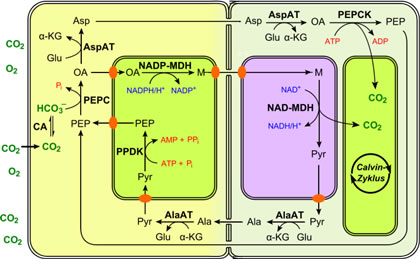
First, carbon dioxide from the atmosphere enters the sugarcane leaf, and fixation is accomplished by the enzyme phosphoenolpyruvate carboxylase (PEP carboxylase). This step takes place within the cytoplasm, not inside the chloroplasts. The first stable product is a four-carbon organic compound that is an acid, usually malate. Sugarcane and other plants with this photosynthetic pathway are known as C plants.
In C4 plants, this photosynthetic pathway is tied to a unique leaf anatomy known as Kranz anatomy. This term refers to the fact that in C4 plants the cells that surround the water- and carbohydrate conducting system (known as the vascular system) are packed very tightly together and are called bundle sheath cells.
Surrounding the bundle sheath is a densely packed layer of mesophyll cells. The densely packed mesophyll cells are in contact with air spaces in the leaf, and because of their dense packing they keep the bundle sheath cells from contact with air. This Kranz anatomy plays a major role in C4 photosynthesis.
In C4 plants the initial fixation of carbon dioxide from the atmosphere takes place in the densely packed mesophyll cells. After the carbon dioxide is fixed into a four-carbon organic acid, the malate is transferred through tiny tubes from these cells to the specialized bundle sheath cells.
Inside the bundle sheath cells, the malate is chemically broken down into a smaller organic molecule, and carbon dioxide is released. This carbon dioxide then enters the chloroplast of the bundle sheath cell and is fixed a second timewith the enzyme Rubisco and continues through the C3 pathway.
Advantages of Double-Carbon Fixation
The double-carbon fixation pathway confers a greater photosynthetic efficiency on C4 plants over C3 plants, because the C3 enzyme Rubisco is highly inefficient in the presence of elevated levels of oxygen. In order for the enzyme to operate, carbon dioxide must first attach to the enzyme at a particular location known as the active site.
However, oxygen is also able to attach to this active site and prevent carbon dioxide from attaching, a process known as photorespiration. As a consequence, there is an ongoing competition between these two gases for attachment at the active site of the Rubisco enzyme. Not only does the oxygen outcompete carbon dioxide; when oxygen binds to Rubisco, it also destroys some of the molecules in the Calvin
cycle.
At any given time, the winner of this competition is largely dictated by the relative concentrations of these two gases. When a plant opens its stomata (the pores in its leaves), the air that diffuses in will be at equilibrium with the atmosphere, which is 21 percent oxygen and 0.04 percent carbon dioxide.
During hot, dry weather, excess water vapor diffuses out, and under these conditions plants face certain desiccation if the stomata are left open continuously.When these pores are closed, the concentration of gases will change. As photosynthesis proceeds, carbon dioxide will be consumed and oxygen generated.
When the concentration of carbon dioxide drops below 0.01 percent, oxygen will outcompete carbon dioxide at the active site, and no net photosynthesis occurs. C4 plants, however, are able to prevent photorespiration, because the PEP carboxylase enzyme is not inhibited by oxygen.
Thus, when the stomata are closed, this enzyme continues to fix carbon inside the leaf until it is consumed. Because the bundle sheath is isolated from the leaf?s air spaces, it is not affected by the rising oxygen levels, and the C3 cycle functions without interference. C4 photosynthesis is found in at least nineteen families of flowering plants.
No family is exclusively composed of C4 plants. Because C4 photosynthesis is an adaptation to hot, dry environments, especially climates found in tropical regions, C4 plants are often able to out compete C3 plants in those areas. In more temperate regions, they have less of an advantage and are therefore less common.
CAM Photosynthesis
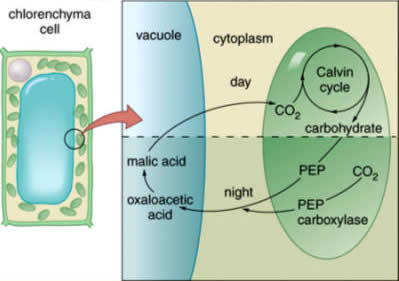
A second alternative photosynthetic pathway, known as crassulacean acid metabolism (CAM), exists in succulents such as cacti and other desert plants. These plants have the same two carbon-fixing steps as are present in C4 plants, but rather than being spatially separated between the mesophyll and bundle sheath cells, CAM plants have both carbon dioxide-fixing enzymes within the same cell.
These enzymes are active at different times, PEP carboxylase during the day and Rubisco at night. Just as Kranz anatomy is unique to C4 plants, CAM plants are unique in that the stomata are open at night and largely closed during the day.
The biochemical pathway of photosynthesis in CAM plants begins at night. With the stomata open, carbon dioxide diffuses into the leaf and into mesophyll cells, where it is fixed by the C4 enzyme PEP carboxylase. The product is malate, as in C4 photosynthesis, but it is transformed into malic acid (a nonionic form of malate) and is stored in the cell?s vacuoles (cavities within the cytoplasm) until the next day.
Although the malic acid will be used as a carbon dioxide source for the C3 cycle, just as in C4 photosynthesis, it is stored until daylight because the C3 cycle requires light as an energy source. The vacuoles will accumulate malic acid through most of the night.
A few hours before daylight, the vacuole will fill up, and malic acid will begin to accumulate in the cytoplasm outside the vacuole. As it does, the pH of the cytoplasm will become acidic, causing the enzyme to stop functioning for the rest of the night.
When the sun rises the stomata will close, and photosynthesis by the C3 cycle will quickly deplete the atmosphere within the leaf of all carbon dioxide. At this time, the malic acid will be transported out of the vacuole to the cytoplasmof the cell. There it will be broken down, and the carbon dioxide will enter the chloroplast and be used by the C3 cycle; thus, photosynthesis is able to continue with closed stomata.
Crassulacean acid metabolism derives its name from the fact that it involves a daily fluctuation in the level of acid within the plant and that it was first discovered to be common in species within the stonecrop family, Crassulaceae.
The discovery of this photosynthetic pathway dates back to the 1960?s. The observation that succulent plants become very acidic at night, however, dates back to at least the seventeenth century, when it was noted that cactus tastes sour in the morning and bitter in the afternoon.
CAM Plant Ecosystems
There are two distinctly different ecological environments where CAM plants may be found. Most are terrestrial plants typical of deserts or other harsh, dry sites.
In these environments, the pattern of stomatal opening and closing provides an important advantage for surviving arid conditions: When the stomata are open, water is lost; however, the rate of loss decreases as the air temperature decreases. By restricting the time period of stomatal opening to the nighttime, CAM plants are extremely good at conserving water.
The other ecological setting where CAM plants are found is in certain aquatic habitats. When this environment was first discovered, it seemed quite odd, because in these environments conserving water would be of little value to a plant. It was found, however, that there are aspects of the aquatic environment which make CAM photosynthesis advantageous.
In shallow bodies of water, the photosynthetic consumption of carbon dioxide may proceed at a rate in excess of the rate of diffusion of carbon dioxide from the atmosphere into the water, largely because gases diffuse several times more slowly in water than in air.
Consequently, pools of water may be completely without carbon dioxide for large parts of the day. Overnight, carbon dioxide is replenished, and aquatic CAM plants take advantage of this condition to fix the plentiful supply of carbon dioxide available at night and store it as malic acid.
Hence, during the day, when the ambient carbon dioxide concentration is zero, these plants have their own internal supply of carbon dioxide for photosynthesis. Thus, two very different ecological conditions have selected for the identical biochemical pathway.
These two modified photosynthetic pathways adequately describe what happens in most terrestrial plants, although there is much variation. For example, there are species that appear in many respects to have photosynthetic characteristics intermediate to C3 and C4 plants.
Other plants are capable of switching from exclusively C3 photosynthesis to CAM photosynthesis at different times of the year. Photosynthesis by aquatic plants appears to present even more variation. C3-C4 intermediate plants seem to be relatively common compared to the terrestrial flora, and several species have C4 photosynthesis but lack Kranz anatomy.
- Calvin Cycle
Calvin cycleThe Calvin cycle is the principal mechanism that leads to the conversion of carbon dioxide into sugars by plants, algae, photosynthetic bacteria, and certain other bacteria that use chemicals as an energy source instead of light. The Calvin...
- Carbon Cycle
The carbon cycle is the movement of the element carbon through the earth?s rock and sediment, the aquatic environment, land environments, and the atmosphere. Large amounts of organic carbon can be found in both living organisms and dead organic material....
- Gas Exchange In Plants
Gas Exchange in PlantsGas exchange is the process whereby water vapor and oxygen leave and carbon dioxide enters plant leaves. The gaseous balance in plants is quite complex because plant cells carry on both respiration and photosynthesis. All living...
- Leaf Anatomy
Leaf Anatomy The leaf has evolved as the chief part of the plant for gathering light energy from the sun and conducting photosynthesis to transform that light energy into biochemical energy. Hence, its structure is adapted to that function. Leaves are...
- Photosynthesis
PhotosynthesisPhotosynthesis is the process by which organic sugar molecules are synthesized from an inorganic carbon source (carbon dioxide or bicarbonate), using sunlight as the energy source to drive the process. Although most often associated with...
Agriculture
C4 and CAM Photosynthesis
Alternative forms of photosynthesis are used by specific types of plants, called C4 and CAM plants, to alleviate problems of photorespiration and excess water loss.
Photosynthesis is the physiological process whereby plants use the sun?s radiant energy to produce organic molecules. The backbone of all such organic compounds is a skeleton composed of carbon atoms. Plants use carbon dioxide from the atmosphere as their carbon source.
The overwhelming majority of plants use a single chemical reaction to attach carbon dioxide from the atmosphere onto an organic compound, a process referred to as carbon fixation. This process takes place inside specialized structures within the cells of green plants known as chloroplasts.
The enzyme that catalyzes this fixation is ribulose bisphosphate carboxylase (Rubisco), and the first stable organic product is a three-carbon molecule. This three-carbon compound is involved in the biochemical pathway known as the Calvin cycle. Plants using carbon fixation are referred to as C3 plants because the first product made with carbon dioxide is a three-carbon molecule.
C4 Photosynthesis
For many years scientists thought that the only way photosynthesis occurred was through C3 photosynthesis. In the early 1960?s, however, researchers studying the sugarcane plant discovered a biochemical pathway that involved incorporation of carbon dioxide into organic products at two different stages.
First, carbon dioxide from the atmosphere enters the sugarcane leaf, and fixation is accomplished by the enzyme phosphoenolpyruvate carboxylase (PEP carboxylase). This step takes place within the cytoplasm, not inside the chloroplasts. The first stable product is a four-carbon organic compound that is an acid, usually malate. Sugarcane and other plants with this photosynthetic pathway are known as C plants.
In C4 plants, this photosynthetic pathway is tied to a unique leaf anatomy known as Kranz anatomy. This term refers to the fact that in C4 plants the cells that surround the water- and carbohydrate conducting system (known as the vascular system) are packed very tightly together and are called bundle sheath cells.
Surrounding the bundle sheath is a densely packed layer of mesophyll cells. The densely packed mesophyll cells are in contact with air spaces in the leaf, and because of their dense packing they keep the bundle sheath cells from contact with air. This Kranz anatomy plays a major role in C4 photosynthesis.
In C4 plants the initial fixation of carbon dioxide from the atmosphere takes place in the densely packed mesophyll cells. After the carbon dioxide is fixed into a four-carbon organic acid, the malate is transferred through tiny tubes from these cells to the specialized bundle sheath cells.
Inside the bundle sheath cells, the malate is chemically broken down into a smaller organic molecule, and carbon dioxide is released. This carbon dioxide then enters the chloroplast of the bundle sheath cell and is fixed a second timewith the enzyme Rubisco and continues through the C3 pathway.
Advantages of Double-Carbon Fixation
The double-carbon fixation pathway confers a greater photosynthetic efficiency on C4 plants over C3 plants, because the C3 enzyme Rubisco is highly inefficient in the presence of elevated levels of oxygen. In order for the enzyme to operate, carbon dioxide must first attach to the enzyme at a particular location known as the active site.
However, oxygen is also able to attach to this active site and prevent carbon dioxide from attaching, a process known as photorespiration. As a consequence, there is an ongoing competition between these two gases for attachment at the active site of the Rubisco enzyme. Not only does the oxygen outcompete carbon dioxide; when oxygen binds to Rubisco, it also destroys some of the molecules in the Calvin
cycle.
At any given time, the winner of this competition is largely dictated by the relative concentrations of these two gases. When a plant opens its stomata (the pores in its leaves), the air that diffuses in will be at equilibrium with the atmosphere, which is 21 percent oxygen and 0.04 percent carbon dioxide.
During hot, dry weather, excess water vapor diffuses out, and under these conditions plants face certain desiccation if the stomata are left open continuously.When these pores are closed, the concentration of gases will change. As photosynthesis proceeds, carbon dioxide will be consumed and oxygen generated.
When the concentration of carbon dioxide drops below 0.01 percent, oxygen will outcompete carbon dioxide at the active site, and no net photosynthesis occurs. C4 plants, however, are able to prevent photorespiration, because the PEP carboxylase enzyme is not inhibited by oxygen.
Thus, when the stomata are closed, this enzyme continues to fix carbon inside the leaf until it is consumed. Because the bundle sheath is isolated from the leaf?s air spaces, it is not affected by the rising oxygen levels, and the C3 cycle functions without interference. C4 photosynthesis is found in at least nineteen families of flowering plants.
No family is exclusively composed of C4 plants. Because C4 photosynthesis is an adaptation to hot, dry environments, especially climates found in tropical regions, C4 plants are often able to out compete C3 plants in those areas. In more temperate regions, they have less of an advantage and are therefore less common.
CAM Photosynthesis
A second alternative photosynthetic pathway, known as crassulacean acid metabolism (CAM), exists in succulents such as cacti and other desert plants. These plants have the same two carbon-fixing steps as are present in C4 plants, but rather than being spatially separated between the mesophyll and bundle sheath cells, CAM plants have both carbon dioxide-fixing enzymes within the same cell.
These enzymes are active at different times, PEP carboxylase during the day and Rubisco at night. Just as Kranz anatomy is unique to C4 plants, CAM plants are unique in that the stomata are open at night and largely closed during the day.
The biochemical pathway of photosynthesis in CAM plants begins at night. With the stomata open, carbon dioxide diffuses into the leaf and into mesophyll cells, where it is fixed by the C4 enzyme PEP carboxylase. The product is malate, as in C4 photosynthesis, but it is transformed into malic acid (a nonionic form of malate) and is stored in the cell?s vacuoles (cavities within the cytoplasm) until the next day.
Although the malic acid will be used as a carbon dioxide source for the C3 cycle, just as in C4 photosynthesis, it is stored until daylight because the C3 cycle requires light as an energy source. The vacuoles will accumulate malic acid through most of the night.
A few hours before daylight, the vacuole will fill up, and malic acid will begin to accumulate in the cytoplasm outside the vacuole. As it does, the pH of the cytoplasm will become acidic, causing the enzyme to stop functioning for the rest of the night.
When the sun rises the stomata will close, and photosynthesis by the C3 cycle will quickly deplete the atmosphere within the leaf of all carbon dioxide. At this time, the malic acid will be transported out of the vacuole to the cytoplasmof the cell. There it will be broken down, and the carbon dioxide will enter the chloroplast and be used by the C3 cycle; thus, photosynthesis is able to continue with closed stomata.
Crassulacean acid metabolism derives its name from the fact that it involves a daily fluctuation in the level of acid within the plant and that it was first discovered to be common in species within the stonecrop family, Crassulaceae.
The discovery of this photosynthetic pathway dates back to the 1960?s. The observation that succulent plants become very acidic at night, however, dates back to at least the seventeenth century, when it was noted that cactus tastes sour in the morning and bitter in the afternoon.
CAM Plant Ecosystems
There are two distinctly different ecological environments where CAM plants may be found. Most are terrestrial plants typical of deserts or other harsh, dry sites.
In these environments, the pattern of stomatal opening and closing provides an important advantage for surviving arid conditions: When the stomata are open, water is lost; however, the rate of loss decreases as the air temperature decreases. By restricting the time period of stomatal opening to the nighttime, CAM plants are extremely good at conserving water.
The other ecological setting where CAM plants are found is in certain aquatic habitats. When this environment was first discovered, it seemed quite odd, because in these environments conserving water would be of little value to a plant. It was found, however, that there are aspects of the aquatic environment which make CAM photosynthesis advantageous.
In shallow bodies of water, the photosynthetic consumption of carbon dioxide may proceed at a rate in excess of the rate of diffusion of carbon dioxide from the atmosphere into the water, largely because gases diffuse several times more slowly in water than in air.
Consequently, pools of water may be completely without carbon dioxide for large parts of the day. Overnight, carbon dioxide is replenished, and aquatic CAM plants take advantage of this condition to fix the plentiful supply of carbon dioxide available at night and store it as malic acid.
Hence, during the day, when the ambient carbon dioxide concentration is zero, these plants have their own internal supply of carbon dioxide for photosynthesis. Thus, two very different ecological conditions have selected for the identical biochemical pathway.
These two modified photosynthetic pathways adequately describe what happens in most terrestrial plants, although there is much variation. For example, there are species that appear in many respects to have photosynthetic characteristics intermediate to C3 and C4 plants.
Other plants are capable of switching from exclusively C3 photosynthesis to CAM photosynthesis at different times of the year. Photosynthesis by aquatic plants appears to present even more variation. C3-C4 intermediate plants seem to be relatively common compared to the terrestrial flora, and several species have C4 photosynthesis but lack Kranz anatomy.
- Calvin Cycle
Calvin cycleThe Calvin cycle is the principal mechanism that leads to the conversion of carbon dioxide into sugars by plants, algae, photosynthetic bacteria, and certain other bacteria that use chemicals as an energy source instead of light. The Calvin...
- Carbon Cycle
The carbon cycle is the movement of the element carbon through the earth?s rock and sediment, the aquatic environment, land environments, and the atmosphere. Large amounts of organic carbon can be found in both living organisms and dead organic material....
- Gas Exchange In Plants
Gas Exchange in PlantsGas exchange is the process whereby water vapor and oxygen leave and carbon dioxide enters plant leaves. The gaseous balance in plants is quite complex because plant cells carry on both respiration and photosynthesis. All living...
- Leaf Anatomy
Leaf Anatomy The leaf has evolved as the chief part of the plant for gathering light energy from the sun and conducting photosynthesis to transform that light energy into biochemical energy. Hence, its structure is adapted to that function. Leaves are...
- Photosynthesis
PhotosynthesisPhotosynthesis is the process by which organic sugar molecules are synthesized from an inorganic carbon source (carbon dioxide or bicarbonate), using sunlight as the energy source to drive the process. Although most often associated with...